

Last Updated: August 3, 2026
Cyclic Nucleotides in Signal Transduction
Cyclic nucleotides represent a family of naturally occurring nucleotide analogs that act as second messengers in hormone and ion channel signal transduction processes. Structurally the cyclic nucleotides are single phosphate nucleotides that harbor a “cyclic” bond between the phosphate and both the 3′-OH and the 5′-OH of the ribose portion of the nucleotide. The major cyclic nucleotides are derived from the purines, adenosine and guanosine forming 3′,5′-cyclic adenosine monophosphate (cAMP) and 3′,5′-cyclic guanosine monophosphate (cGMP), respectively. However, the existence of the cyclic pyrimidines [3’,5’-cyclic uridine monophosphate (cUMP) and 3’,5’-cyclic cytidine monophosphate (cCMP)] has been documented in human cells and the degradation of cUMP has been demonstrated to be catalyzed by one of the known cyclic nucleotide phosphodiesterases.
The production and utilization of cAMP and cGMP is paramount for the proper regulation of numerous biochemical and physiological processes. The primary effects of cAMP and cGMP are exerted through the activation of their respective cyclic nucleotide dependent kinases, PKA and PKG.
Additional important effectors of cAMP are the exchange proteins activated by cAMP, EPAC. EPAC proteins are guanine-nucleotide exchange factor (GEF) proteins that are directly activated by cAMP. Another family of proteins that are effectors of cAMP are the Popeye domain containing (Popdc) proteins. The Popdc proteins play a role in tight junction formation and epithelial cell functions.
The cyclic nucleotides themselves play important roles directly by, for example, modulating the activity of ligand-gated ion channels.
Once produced, both cAMP and cGMP (as well as the cyclic pyrimidines) must be degraded to limit the extent of the activities. The degradation of cyclic nucleotides is catalyzed by cyclic nucleotide-specific phosphodiesterases which are covered in the Cyclic Nucleotide Phosphodiesterases section.
Adenylate Cyclases
The adenylate cyclases (adenylyl cyclases) are a family of enzymes that generate the second messenger molecule, cyclic adenosine-3′,5′-monophosphate (cAMP) from ATP. Adenylate cyclases (AC) belong to the family of nucleotidyl cyclase enzymes that is divided into six broad classes identified as Class I–Class VI. All of the adenylate cyclase enzymes expressed in humans belong to the class III family. The human adenylate cyclases are further divided into two distinct sub-types, the transmembrane adenylate cyclases (tmAC) and the soluble adenylate cyclase (sAC). Humans express ten adenylate cyclase genes with nine of the genes (ADCY1–ADCY9) encoding tmAC isoforms and one gene (ADCY10) encoding a sAC isoform.
Transmembrane Adenylate Cyclases
The transmembrane localized adenylate cyclases possess two sets of six transmembrane segments separated by two cytoplasmic segments. The two sets of transmembrane segments are designated TM1-TM6 and TM7-TM12. The two cytoplasmic segments are designated C1 and C2 (or C1 and C2). These cytoplasmic domains are commonly referred to as cyclase homology domains (CHD). The two cytoplasmic domains associate with each other forming a heterodimeric catalytic core of the enzyme. The C1 domain is located C-terminal to the TM6 domain and the C2 domain is located C-terminal to the TM12 domain. Adenylate cyclase isoforms have been characterized that result from alternative splicing events and which contain a single CHD. The CHD in these isoforms homodimerizes to form the catalytic core.
The catalytic domains of the AC are divided into subdomains identified as C1a, C1b, C2a, and C2b. The C1a domain is linked to TM6 while the C2a domain is linked to TM12. These interactions form what is referred to as the helical domain. The actual catalytic site of each AC resides at the interface between the C1a and C2a domains. The C1b and C2b domains are involved in the regulation of AC activity and control isoform specific regulatory functions.
Regulation of tmAC Activity
All of the tmAC enzymes are activated by the alpha-(α)subunit of Gs-type (Gαs) heterotrimeric G-proteins. Inhibition of several tmAC enzymes can be effected by the α-subunit of Gi-type (Gαi) heterotrimeric G-proteins. The Gαi subunits are known to inhibit AC1, AC5, and AC6. The interaction of the Gαs subunit with the C2a domain of tmAC results in the activation of adenylate cyclase activity whereas the interaction of the Gαi subunit with the C1a domain results in inhibition of adenylate cyclase activity.
Calcium ions influence the activity of several of the tmAC either positively (AC1, AC3, and AC8) or negatively (AC5 and AC6). The activation of tmAC activity by Ca2+ involves the calcium-binding regulatory subunit, calmodulin. The mechanism by which Ca2+ inhibits tmAC activity is by inducing the displacement of the Mg2+ cofactor.
The activity of various tmAC isoforms are also regulated by PKA or PKC. PKA inhibits the activity of AC6 and AC8. PKC activates AC1, AC2, AC3, AC5 and AC7, but inhibits AC6.
Nitric oxide (NO) has also been shown to inhibit the activity of AC5 and AC6.
Soluble Adenylate Cyclase
The soluble adenylate cyclase is not localized to the cytosol but is actually distributed within discrete sub-domains within the cell such as the nucleus, mitochondria, mitotic spindle, and cilia. The discrete localization allows for, what are referred to as, microdomains of cAMP production and action to be established. Soluble AC is tethered to these sites by being anchored to PKA or scaffold proteins called A-kinase anchoring proteins (AKAP). Like the tmAC enzymes, the sAC enzyme also possesses two domains (also identified as C1 and C2) that associate together to form the catalytic core of the enzyme.
Unlike the tmAC, sAC activity is not regulated by heterotrimeric G-proteins. The activity of sAC is enhanced by both bicarbonate (HCO3–) and Ca2+. The effect of bicarbonate is to enhance substrate (ATP) turnover and reduce substrate inhibition, while Ca2+ enhances substrate binding. The sAC isoform can also be activated by the presence of another divalent cation, Mg2+ similarly to the role of this divalent cation in the regulation of several of the tmAC isoforms. The ability of HCO3– to regulate sAC activity allows for the enzyme to serve as a sensor for the CO2/HCO3– ratio and thus, indirectly as a pH sensor.
Human sAC plays important roles in airway epithelial cells, the exocrine pancreas, renal epithelium, skin, eyes, and the nervous system. Within the nervous system, sAC is involved as a modulator of chemoreceptor sensation in the carotid and aortic bodies and the brain stem that regulate respiratory rate.
Table of Adenylate Cyclases Expressed in Humans
| Adenylate Cyclase | Gene | Location/ Structure | Comments |
| AC1 | ADCY1 | 7p12.3: 22 exons | primarily expressed in the brain, retina, and the adrenal medulla; stimulated by Ca2+ (via calmodulin) and PKA-mediated phosphorylation; inhibited by Gαi and by Gβγ dimers; involved in regulation of circadian rhythms likely by modulating cAMP production in the retina; pathogenic variants in ADCY1 associated with autosomal recessive deafness type 44 (DFNB44) |
| AC2 | ADCY2 | 5p15.3: 25 exons | highest levels of expression seen in brain, skeletal muscle, and lung; stimulated by Gβγ dimers following Gαs activation; activity enhanced by PKA phosphorylation |
| AC3 | ADCY3 | 2p23.3: 26 exons | highest expression seen in pancreas, olfactory epithelium, brain, brown adipose tissue (BAT), also expressed in lung, heart, brain, kidney, and liver; activated by Ca2+ (via calmodulin) and PKA; inhibited by Gαi |
| AC4 | ADCY4 | 14q12: 26 exons | highest expression seen in brain but also widely expressed; stimulated by Gαs and by Gβγ dimers and by PKA |
| AC5 | ADCY5 | 3q21.1: 29 exons | highest levels of expression in cardiac myocytes and brain striatum; inhibited by Gαi, Ca2+, and PKA-mediated phosphorylation; stimulated by Gβγ complexes formed from the proteins encoded by the GNB1 and GNG2 genes; pathogenic variants in the ADCY5 gene associated with familial dyskinesia with facial myokymia (FDFM) |
| AC6 | ADCY6 | 12q12–q13: 23 exons | widely expressed; functions in β-adrenergic receptor-stimulated signaling in cardiac myocytes and vascular smooth muscle (VSM), also functions in vasopressin-mediated water resorption in the kidney; inhibited by Gαi, PKA-mediated phosphorylation, Ca2+, and NO; stimulated by Gβγ complexes formed from the proteins encoded by the GNB1 and GNG2 genes |
| AC7 | ADCY7 | 16q12.1: 31 exons | highest expression in brain and platelets but also widely expressed; stimulated by Gβγ dimers and by PKA-mediated phosphorylation |
| AC8 | ADCY8 | 8q24: 18 exons | highest levels of expression seen in brain and lung; inhibited by Gαi; stimulated by Ca2+ (via calmodulin); plays a role in learning, memory, and drug dependence |
| AC9 | ADCY9 | 16p13.3: 12 exons | highest expression in airway smooth muscle, also expressed in lung, pituitary and skeletal muscle; inhibited by Gαi; stimulated by β-adrenergic receptors; may be a membrane docking site for calcineurin |
| AC10 | ADC10 | 1q24: 36 exons | soluble enzyme (sAC); expressed at highest levels in testes but also widely expressed; insensitive to G-protein regulation; regulated by bicarbonate ion and Ca2+; alternative mRNA splicing yields at least three distinct sAC isoforms, two of the alternatively spliced mRNAs generate isoforms of sAC that contain only one (C2 only) of the two catalytic domains |
The PKA Family of Kinases
Once produced by the action of an adenylate cyclase, cAMP exerts its effects in a number of ways, either directly or by interaction with the regulatory subunits of cAMP-dependent protein kinases (PKA). In the absence of cAMP, PKA enzymes exist as inactive heterotetrameric complexes composed of two regulatory and two catalytic subunits.
Humans express three different catalytic subunit genes, PRKACA (protein kinase cAMP-activated catalytic subunit alpha), PRKACB (protein kinase cAMP-activated catalytic subunit beta), PRKACG (protein kinase cAMP-activated catalytic subunit gamma).
Humans express four different PKA regulatory subunit genes, PRKAR1A (protein kinase cAMP-dependent type I regulatory subunit alpha), PRKAR1B (protein kinase cAMP-dependent type I regulatory subunit beta), PRKAR2A (protein kinase cAMP-dependent type II regulatory subunit alpha), and PRKAR2B (protein kinase cAMP-dependent type II regulatory subunit beta). These four genes encode four different proteins that exhibit tissue-specific patterns of expression and exert distinct roles in cell differentiation and growth control.
The original model proposed for the role of cAMP in the regulation of PKA suggested that two molecules of cAMP bind to each of the two regulatory subunits of PKA stimulating the release of the two catalytic subunits. Once released the catalytic subunits were free to phosphorylate serine and/or threonine residues in numerous target proteins. However, recent experimental data has found that active PKA remains in the heterotetrameric form and that cAMP functions, not to completely displace the catalytic subunits, but to facilitate the movement of the catalytic sites of the catalytic subunits away from the inhibitory domain of the regulatory subunits allowing the catalytic subunits to interact with, and phosphorylate, substrate proteins.
The activity of a given PKA isoform is ultimately regulated by its localization, such as cytosolic versus nuclear. Localization of PKA is effected by interaction with proteins of the A-kinase-anchoring protein (AKAP) family. Humans express 31 genes that encode proteins of the AKAP family. Proteins of the AKAP family that function in the localization of PKA to discrete subcellular locations do so by interaction with the regulatory subunits of the enzymes. The AKAP proteins also serve as docking sites for other proteins that contribute to the overall regulation of PKA activity such as phosphatases and phosphodiesterases. Indeed, the activity of PKA is self-limiting due to the fact that one of the substrates for the enzyme is a member of the cyclic nucleotide phosphodiesterase (PDE) family (see section below for details on PDE) that when active hydrolyzes cAMP to AMP.
PKA-Independent Effects of cAMP
In addition to serving to activate the catalytic activity of PKA, cAMP is capable of exerting numerous effects independent of PKA. Several of these effects are the result of cAMP interacting with guanine nucleotide exchange factors (GEF) that includes EPAC1 (Exchange Protein directly Activated by cAMP 1) and EPAC2. These cAMP interacting proteins are encoded by the RAPGEF3 (RAP guanine nucleotide exchange factor 3) and RAPGEF4 genes, respectively.
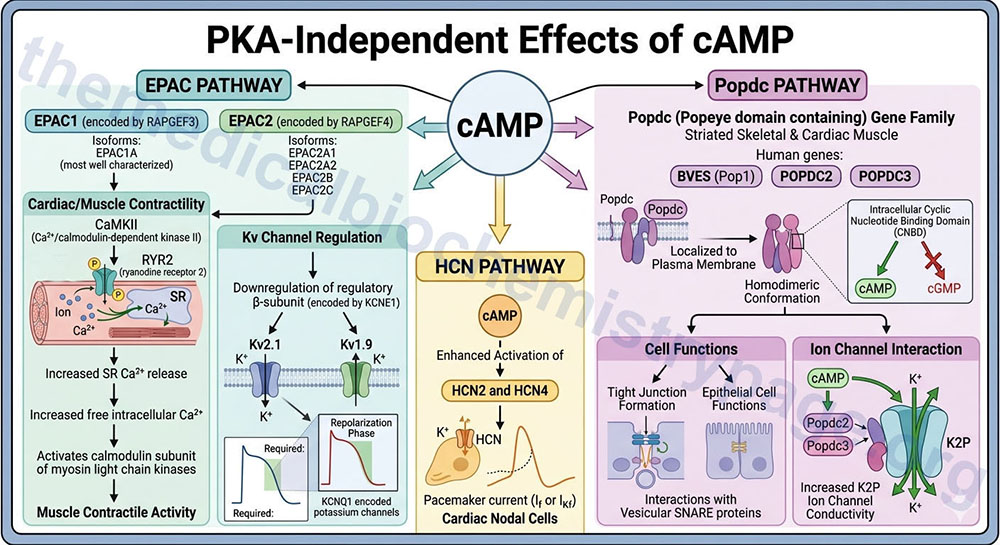
Alternative splicing of RAPGEF3 results in two protein isoforms with the larger isoform identified as EPAC1A being the most well characterized. Alternative splicing and alternative promoter utilization increases the complexity of RAPGEF4 isoforms such that at least four isoforms have currently been well characterized; EPAC2A1, EPAC2A2, EPAC2B, and EPAC2C.
Many direct effects of cAMP are exerted within the peripheral nervous system such as in the autonomic control of the cardiovascular system. The activation of EPAC1 by cAMP results in the activation of Ca2+/calmodulin-dependent kinase II (CaMKII). When activated, CaMKII phosphorylates the calcium release channel, ryanodine receptor 2, RYR2, resulting in increased release of Ca2+ stored within the sarcoplasmic reticulum, SR. Increased free intracellular Ca2+ plays a major role in the contractile activity of muscle cells through, among other effects, the activation of the calmodulin subunit of myosin light chain kinases.
Activation of EPAC also results in the downregulation of the regulatory subunit of voltage-gated potassium channels which are commonly identified as Kv channels. The regulatory subunit of the Kv channels is encoded by the KCNE1 gene and is commonly referred to as the β-subunit. The specific cardiac Kv family channels that are regulated by the KCNE1 encoded protein are Kv2.1 channels encoded by the KCNB1 gene and Kv1.9 channels encoded by the KCNQ1 gene. The KCNQ1 encoded potassium channels are required for the repolarization phase of cardiac myocyte action potentials.
In addition to cAMP effects on the EPAC proteins, cAMP binds to, and enhances the activation of, the hyperpolarization-activated cyclic nucleotide-gated channel 2 and 4 (HCN2 and HCN4) that are responsible for the pacemaker current (If or IKf) in cardiac nodal cells.
Another family of cAMP-binding proteins is the Popeye domain containing (Popdc) gene family. This family is so-called based on the observation that the original family member, Popeye1 (Pop1), was highly expressed in striated skeletal muscle. Humans express three genes encoding proteins in this family identified as BVES, POPDC2, and POPDC3. The BVES (blood vessel epicardial substance), which encodes Pop1 is the name given to the gene by one of two groups that simultaneously identified the founding member of the Popeye domain containing family. All three genes in this family are expressed in cardiac and skeletal muscle.
All three Popdc proteins are localized to the plasma membrane where they exist in homodimeric conformation. The intracellular portion of the Popdc proteins harbors a cyclic nucleotide binding domain (CNBD) that allows for binding of cAMP, but this domain in the Popdc proteins does not bind cGMP. The Popdc proteins play a role in tight junction formation and epithelial cell functions, in part through interactions with vesicular SNARE proteins. Popdc proteins also interact with ion channels, specifically members of the two-pore domain potassium channel (K2P) family. The binding of cAMP to Popdc2 and Popdc3 results in an increase in ion channel conductivity.
Guanylate Cyclases
The guanylate cyclases are a family of enzymes that generate the second messenger molecule, cyclic guanosine-3′,5′-monophosphate (cGMP) from GTP. Like cAMP, cGMP acts as a second messenger molecule in a variety of signal transduction events. Intracellular cGMP has been shown to play a critical role in the modulation of lipolysis, platelet aggregation, blood pressure, gut peristalsis, intestinal fluid secretion, neurotransmission, vision, sexual arousal, long bone growth, cardiac myocyte growth, and oocyte maturation. The guanylate cyclases are regulated by a broad spectrum of agents that includes nitric oxide (NO), bicarbonate ion (HCO3–), the natriuretic peptides (ANP, BNP, and CNP), and the guanylyl cyclase activating proteins (GCAP).
The guanylate cyclase (GC) enzymes, like the adenylate cyclase enzymes, belong to the class III family of nucleotidyl cyclases. There are two distinct types of guanylate cyclases expressed in humans, the soluble enzymes (sGC) and the sub-family of single transmembrane-spanning guanylate cyclases.
The sGC family members all function as heterodimers of one α-subunit and one β-subunit. The heme moiety, to which NO binds, is attached to the N-terminus of the β-subunits. The catalytic activity of the sGC enzymes is created by C-terminal domains in both subunits.
The human α1 subunit (also identified as α3) is encoded by the GUCY1A1 gene (also identified as GUCY1A3). The GUCY1A1 gene is located on chromosome 4q32.1 and is composed of 13 exons that generate 18 alternatively spliced mRNAs, 14 of which encode the same protein identified as isoform A (690 amino acids).
The α2 protein is encoded by the GUCY1A2 gene. The GUCY1A2 gene is located on chromosome 11q22.3 and is composed of 9 exons that generate two alternatively spliced mRNAs encoding isoform 1 (763 amino acids) and isoform 2 (732 amino acids). The α2 subunit of sGC heterodimers may actually direct this isoform of sGC to membranes. The highest concentrations of the α2/β1 isoform of sGC is in the brain with the α1/β1 isoform predominating in all other tissues.
The human genome contains two β-subunit genes but one (GUCY1B2) is a pseudogene. The β1 subunit (also identifies as β3) is encoded by the GUCY1B1 gene (also identified as GUCY1B3). The GUCY1B1 gene is located on chromosome 4q32.1 and is composed of 17 exons that generate six alternatively spliced mRNAs, each of which encode a distinct protein isoform.
In addition to the classic role of sGC in the formation of cGMP, recent evidence has shown that the α1β1 enzyme can form cyclic pyrimidine nucleotides (cCMP and cUMP) in the presence of Mn2+, whereas in the presence of Mg2+ only cGMP is formed. As yet, the proteins to which the cyclic pyrimidines bind have not been identified.
Although there are seven transmembrane-spanning guanylate cyclase genes in mammals, humans express only five isoforms designated as GC-A, GC-B, GC-C, GC-E, and GC-F. The GC-D and GC-G encoding genes in humans are pseudogenes. The transmembrane-spanning GC isoforms all serve as receptors for specific ligands. The GC-A and GC-B isoforms are more commonly referred to as the natriuretic peptide receptor 1 (NPR1 or NPR-A) and natriuretic peptide receptor 2 (NPR2 or NPR-B) isoforms. NPR1 binds both ANP and BNP, whereas, NPR2 binds CNP.
All of the membrane-spanning GC isoforms function as homodimers. NPR1 is highly expressed in kidney, lung, adrenal glands, vasculature tissues, brain, liver, endothelial cells, and adipose tissues with lower levels of expression observed in cardiac tissues. When the NPR1 gene is knocked out in mice they develop cardiac hypertrophy, ventricular fibrosis, and hypertension. The activity of the NPR1 protein is regulated by its state of intracellular phosphorylation. The receptor is unable to respond to ligand binding in the dephosphorylated state, thus, requiring phosphorylation for activity.
Guanylate cyclase C (GC-C, encoded by the GUCY2C gene) is a receptor for several endogenous and exogenous peptides. Pathogenic bacteria produce a heat-stable enterotoxin that binds and activates the GC-C protein. When the bacterial enterotoxin binds to the GUCY2C-encoded guanylate cyclase, the increased production of intracellular cGMP results in activation of a cGMP-dependent protein kinase, PKG (specifically PKGII). Under these conditions, PKGII phosphorylates the cystic fibrosis transmembrane conductance regulator (CFTR) resulting in increased Cl– secretion in the gut. As the intestinal ion secretion increases there is concomitant loss of water to the lumen of the gut. The loss of intestinal water leads to severe dehydration and the increased water in the lumen leads to severe diarrhea.
Guanylin and uroguanylin are endogenous peptide ligands for the GUCY2C-encoded guanylate cyclase. Humans express two transmembrane guanylate cyclases in the retina, both of which are involved in the processes of phototransduction. These two enzymes are GC-E (more commonly called retinal guanylate cyclase 1, RETGC-1) and GC-F (more commonly called RETGC-2). The activities of the two RETGC enzymes are regulated, not by ligand binding, but through the action of a GCAP (details below the following Table).
All of the sGC enzymes function as heterodimers. The sGC isoforms possess a heme-binding domain in the N-terminal region of the proteins that is required for activation. The primary endogenous activator of the sGC enzymes is nitric oxide (NO), but carbon monoxide (CO) can also bind to the heme prosthetic group. The soluble GC enzymes are expressed in most human tissues with enriched expression seen in brain, kidney, vascular tissues, and lung.
Although there are four different sGC isoforms that can form heterodimers, the most well characterized forms in human tissues are the α1/β1 and α2/β1 forms. Within these heterodimers it is the β-subunit that contains a His residue in its N-terminal domain that coordinates the ferrous (Fe2+) iron within the heme prosthetic group. Activation of sGC isoforms, by NO, requires the presence of the ferrous iron-heme prosthetic group. There is, however, complex interplay between the α-subunits and the β-subunits in sGC heterodimers that results in activity specificity. For example, disruption of the α1-subunit abolishes NO-dependent platelet aggregation yet there remains a normal vasorelaxation response. Conversely, disruption of the β1-subunit results in loss of both responses.
Table of Single Transmembrane-Spanning Guanylate Cyclases Expressed in Humans
| GC Designation | Gene | Location/ Structure | Comments |
| GC-A | NPR1 | 1q21.3: 22 exons | also identified as GUCY2A and ANPRA; receptor for ANP and BNP; expression in lung, kidney, adrenal gland, vascular smooth muscle, endothelium, heart, adipose tissue |
| GC-B | NPR2 | 9p13.3: 22 exons | also identified as GUCY2B and ANPRB; receptor for CNP; expression in bone, vascular smooth muscle, lung, brain, heart, liver, uterus, and follicle |
| GC-C | GUCY2C | 12p12.3: 27 exons | receptor for the gut-derived peptides, guanylin and uroguanylin; originally identified as the receptor for bacterial heat-stable enterotoxin; highest expression is in apical membrane of intestinal epithelial cells |
| GC-E | GUCY2D | 17p13.1: 20 exons | expressed in the retina; also called retinal guanylate cyclase 1 (RETGC-1) or just guanylate cyclase 1 (GC1); regulates phototransduction in the dark; no ligand, activated by guanylate cyclase activating protein 1 (GCAP1) and GCAP2; pathogenic variants in gene are the cause of Leber congenital amaurosis and autosomal dominant cone rod dystrophy 6 (CRD6) |
| GC-F | GUCY2F | Xq22.3–q23: 20 exons | expressed in the retina; also called retinal guanylate cyclase 2 (RETGC-2) or just guanylate cyclase 2 (GC2); regulates phototransduction in the dark; no ligand, activated by guanylate cyclase activating protein 1 (GCAP1) and GCAP2; pathogenic variants in gene are the causes of X-linked retinitis pigmentosa |
The PKG Family of Kinases
Like the role of cAMP in modulating the activity of PKA, cGMP modulates the activity of the cGMP-dependent protein kinases, PKG. Humans express three distinct isoforms of PKG identified as PKGIα, PKGIβ, and PKGII. Both PKGIα and PKGIβ are soluble enzymes and both are derived from the same gene (PRKG1) as a result of alternative splicing. The PKGII enzyme is a membrane-bound version of the PKG kinase family and is encoded by a separate gene, PRKG2.
The PRKG1 gene is located on chromosome 10q11.23–q21.1 and is composed of 21 exons that generate four alternatively spliced mRNAs.
The PRKG2 gene is located on chromosome 4q21.21 and is composed of 23 exons that generate seven alternatively spliced mRNAs.
Unlike the subunit structure of PKA, PKG enzymes do not possess separate catalytic and regulatory subunits. Active PKG isoforms are homodimers and are activated by direct binding of cGMP to each subunit in the homodimer.
The PKGI isoforms are most highly expressed in vascular smooth muscle cells and in platelets. PKGII is most highly expressed in retinal cells, zona glomerulosa cells in the adrenal cortex, intestinal mucosal cells, and pancreatic duct cells.
Gyanylate Cyclase Activating Proteins, GCAP
Guanylate cyclase activator proteins (GCAP) are a family of proteins that regulate the activity of several members of the single transmembrane-spanning guanylate cyclases. Humans express three GCAP proteins that function in the regulation of retinal cone and rod cell guanylate cyclases. The retinal cell guanylate cyclases are called RetGC-1 (often just simply referred to as GC1) and RetGC-2 (often just simply referred to as GC2).
Humans also express two additional genes encoding proteins of the GCAP family. However, in the case of these latter two genes the encoded proteins (guanylin and uroguanylin) are the ligands for the intestinal transmembrane guanylate cyclase encoded by the GUCY2C gene.
Retinal cells of the eye can adjust their sensitivity to photons as a means to allow for phototransduction over a wide range of light intensities. These adjustments to sensitivity are brought about through the effect that calcium ions (Ca2+) exert on the activity of the retinal cell membrane guanylate cyclases which produce cGMP. For more details on the role of cGMP in phototransduction go to the Vitamins: Water and Fat Soluble page.
The effects of Ca2+ on retinal guanylate cyclase activity is due to regulated function, not of the guanylate cyclases directly, but of guanylate cyclase activating proteins, GCAP. Humans express three GCAP genes, whose encoded proteins are identified as GCAP1, GCAP2, and GCAP3, that function in the regulation of cGMP-mediated phototransduction. Both GCAP1 and GCAP2 bind Ca2+ and have similar dependence and cooperativity for guanylate cyclase activation by Ca2+. The GCAP1 protein is encoded by the GUCA1A gene, the GCAP2 protein is encoded by the GUCA1B gene, and the GCAP3 protein is encoded by the GUCA1C gene.
Guanylin and Uroguanylin Peptides
The guanylin family of intestinal peptides was originally characterized due to attempts to understand how ingested sodium led to increased renal electrolyte and water secretion. These original observations predicted the existence of intestinal natriuretic-like hormones. Following their full characterization these intestinal hormones were called guanylins. Humans express two intestinal guanylin family peptides identified as guanylin and uroguanylin.
Guanylin is a 15 amino acid peptide that contains two intramolecular disulfide bonds, whereas, uroguanylin is a 19 amino acid peptide whose disulfide bonds are conserved relative to those in guanylin. The guanylin peptide is encoded by the GUCA2A gene and uroguanylin is encoded by the GUCA2B. Both guanylin and uroguanylin have been called guanylate cyclase activating proteins (GCAP) with guanylin identified as GCAP2A and uroguanylin identified as GCAP2B.
The GUCA2A gene is located on chromosome 1p34.2 and is composed of 3 exons that encode a 115 amino acid preproprotein.
The GUCA2B gene is located very close to the GUCA2A gene on chromosome 1p34.2 and is also composed of 3 exons that encode a 112 amino acid preproprotein.
In addition to guanylin and uroguanylin, there are two additional guanylin peptide family members identified as lymphoguanylin and renoguanylin. However, as yet a human isoform of renoguanylin remains to be identified.
Following the ingestion of a salty meal, intestinal cells secrete guanylin and uroguanylin into the intestinal lumen where they bind to the GUCY2C-encoded transmembrane guanylate cyclase. Guanylin producing cells of the intestine are goblet and epithelial cells of the colonic mucosa, whereas, uroguanylin producing cells are enterochromaffin cells of the small intestine.
Activation of cGMP production by the intestinal GUCY2C encoded enzyme results in inhibition of sodium absorption from intestinal lumen by inhibiting a Na+/H+ exchange (NHE) transporter (specifically NHE2 encoded by the SLC9A2 gene), increased bicarbonate (HCO3–) and Cl– secretion, inhibited water absorption, and increased renal Na+ and K+ secretion.
These effects of guanylin and uroguanylin, exerted as a result of increased intracellular cGMP production, are exerted both at the level of increased PKG and PKA activity. The increased PKG (specifically PKGII) activity is a direct result of cGMP activating this kinase. PKA activity is also enhanced because the increased cGMP leads to inhibition of phosphodiesterase 3, PDE3, family (see next section) enzyme activity. PDE3 enzymes normally hydrolyze cAMP leading to reduced levels of active PKA. However, with cGMP-mediated inhibition of PDE3 the level of active PKA remains high.
One of the targets for intestinal PKGII is the CFTR protein. Phosphorylation of CFTR by PKGII results in increased Cl– transport into the lumen of the gut. PKA can also phosphorylate the CFTR protein and also phosphorylates an apical membrane HCO3–/Cl– antiporter. The inhibition of the NHE2 transporter occurs directly via cGMP binding to the transporter.
In addition to guanylin peptide secretion into the intestinal lumen, intestinal cells transport guanylin and uroguanylin into the blood. The secretion of these peptides into the blood allows them to interact with their receptor guanylate cyclase in renal tubular cells where the response is natriuresis, diuresis, and kaliuresis. In addition to the intestine, the guanylin and uroguanylin genes are expressed in the pancreas, adrenal glands, lung, and reproductive systems where these peptides exert regulation on membrane transport systems.
Cyclic Nucleotide Phosphodiesterases: PDE
Phosphodiesterases are enzymes that catalyze the hydrolysis of a phosphodiester bond. Humans express a large number of phosphodiesterases, with the cyclic nucleotide phosphodiesterases (PDE) being the class of enzymes most commonly associated with the term, phosphodiesterase. Other mammalian phosphodiesterases include several members of the phospholipase family of enzymes such as the phospholipase D (PLD) enzymes and the phospholipase C (PLC) enzymes. Ribonucleases (RNases) and deoxyribonucleases (DNases) are also members of the phosphodiesterase superfamily of enzymes.
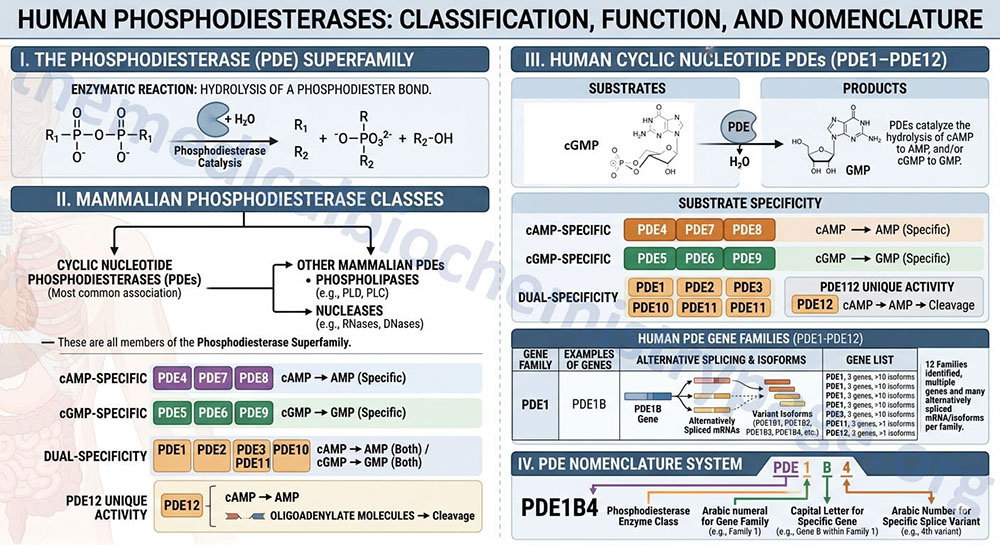
The PDE expressed in human tissues hydrolyze cAMP to AMP, and/or cGMP to GMP. Certain members of the human PDE family are specific for one cyclic nucleotide or the other while some do not exhibit substrate specificity and can hydrolyze either cAMP or cGMP. Humans express 12 PDE gene families identified as PDE1–PDE12. Within each gene family there may be several distinct genes and many of these genes generate alternatively spliced mRNAs encoding variant isoforms of the enzyme.
The cAMP-specific phosphodiesterases are encoded by the PDE4, PDE7, and PDE8 gene families.
The cGMP-specific phosphodiesterases are encoded by the PDE5, PDE6, and PDE9 gene families.
The dual-specificity phosphodiesterases are encoded by the PDE1, PDE2, PDE3, PDE10, and PDE11 gene families.
The activity of PDE12 is unique in that is hydrolyzes cAMP as well as oliogadenylate molecules. The designation for a particular phosphodiesterase gene includes the PDE term followed by an Arabic numeral for the gene family, followed by a capital letter to designate the particular gene in that family. If a gene generates splice variants then there is another Arabic number added to the enzyme nomenclature to designate the specific splice variant. For example, the phosphodiesterase designated PDE1B4 is derived from the fourth splice variant of the PDE1B gene, where the PDE1B gene is one of several family 1 PDE genes.
Table of Mammalian Cyclic Nucleotide Phosphodiesterases (PDEs)
| PDE Gene Family | Family Members | Comments |
| 1 | PDE1A, PDE1B, PDE1C | all members of the PDE1 family are calmodulin-regulated enzymes; PDE1A produces five splice variant isoforms; PDE1B produces four splice variant isoforms; although dual-specific the PDE1B isoforms show preference for cGMP as substrate |
| 2 | PDE2A | generates four splice variants |
| 3 | PDE3A, PDE3B | PDE3 genes are members of the cGMP-inhibited cyclic nucleotide phosphodiesterase (cGI-PDE) family; PDE3A expresses two splice variants; PDE3B involved in insulin-mediated signal transduction events that result in reduced levels of active PKA |
| 4 | PDE4A, PDE4B, PDE4C, PDE4D | PDE4A produces five splice variant isoforms; PDE4B produces six splice variant isoforms; altered activity of PDE4B isoforms are associated with bipolar disorder and schizophrenia; PDE4C produces three splice variant isoforms; PDE4D produces nine spice variant isoforms |
| 5 | PDE5A | PDE5A produces three splice variant isoforms, although PDE5A1 and PDE5A2 only isoforms for which activity has been demonstrated; is critical enzyme involved in smooth muscle cell relaxation in the cardiovascular system; is the target of the PDE5 inhibitor class of drugs used to treat erectile dysfunction and pulmonary hypertension |
| 6 | PDE6A, PDE6B, PDE6C, PDE6D | PDE6A and PDE6B encode the alpha (α) and beta (β) subunits, respectively, of the catalytic core of the rod outer segment phosphodiesterase 6 holoenzyme; the rod PDE6 enzyme is a heterotetrameric complex composed of one α-subunit, one β-subunit, and two γ-subunits (inhibitory subunits); pathogenic variants in PDE6A cause a form of autosomal dominant retinitis pigmentosa; PDE6B produces three splice variant isoforms; pathogenic variants in PDE6B are associated with retinitis pigmentosa and autosomal dominant congenital stationary night blindness; PDE6C encodes the alpha prime (α’) subunit of the cone cell phosphodiesterase 6 which is a complex composed of two α’-subunits and three additional small proteins of 11kDa, 13kDa, and 15kDa; pathogenic variants in PDE6C cause cone dystrophy type 4 (COD4); PDE6D encodes the delta (δ) subunit of a retinal rod phosphodiesterase; PDE6D produces two splice variant isoforms; most PDE5 inhibitors also inhibit rod PDE6 which has significance for contraindication of that class of drug in certain individuals |
| 7 | PDE7A, PDE7B | PDE7A produces two splice variant isoforms; inhibitors of PDE7 isoforms are being tested for efficacy in inflammatory airway disorders and inflammatory disorders in the CNS |
| 8 | PDE8A, PDE8B | PDE8A produces three splice variant isoforms; PDE8B produces five splice variant isoforms; PDE8A1 isoform is involved in activated lymphocyte chemotaxis; pathogenic variants in PDE8B result in autosomal dominant striatal degeneration (ADSD) |
| 9 | PDE9A | PDE9A produces fifteen splice variant isoforms; pharmacologic inhibition of PDE9A may promote cognitive function in patients with Alzheimer disease |
| 10 | PDE10A | PDE10A produces two splice variant isoforms; pharmacologic inhibition of PDE10A is being investigated for the treatment of schizophrenia and Huntington disease |
| 11 | PDE11A | PDE11A produces four splice variant isoforms; pathogenic variants in PDE11A associated with Cushing disease and adrenocortical hyperplasia |
| 12 | PDE12 | originally identified as 2′,5′-phosphodiesterase 12 |